Prevención de la endogamia
[3] Un estudio de 2007 demostró que los ratones endogámicos presentaban una supervivencia significativamente menor cuando se reintroducían en un hábitat natural.
La fecundación cruzada entre individuos no emparentados suele enmascarar los alelos recesivos deletéreos en la progenie.
[8][9] Muchos estudios han demostrado que los individuos homocigóticos suelen estar en desventaja con respecto a los heterocigóticos.
[12] Los mecanismos para evitar la endogamia han evolucionado en respuesta a la selección contra la descendencia consanguínea.
En la naturaleza, los mecanismos para evitar la endogamia son al menos cuatro: reconocimiento de parentesco, dispersión, cópulas extrapareja/extragrupo y retraso en la maduración/supresión reproductiva.
[3][12] Estos mecanismos no se excluyen mutuamente y en una población puede darse más de uno en un momento dado.
El reconocimiento del parentesco es el mecanismo por el que los individuos identifican y evitan aparearse con congéneres estrechamente emparentados.
Existen numerosos ejemplos documentados de casos en los que los individuos consideran poco atractivos a los congéneres estrechamente emparentados.
Los individuos también pueden utilizar sus propias características o fenotipo como plantilla en el reconocimiento de parentesco.
[23] Los resultados indicaron que el olfato puede ayudar a mediar en el desarrollo de la evitación del incesto durante la infancia (el efecto Westermarck).
Se realizaron experimentos con la planta dioica Silene latifolia para comprobar si la selección postpolinización favorece a los donantes de polen menos emparentados y reduce la endogamia.
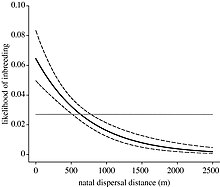
Algunas especies adoptan la dispersión como una forma de separar a los parientes cercanos y evitar la endogamia.
Los desplazamientos a larga distancia pueden conllevar riesgos de mortalidad y costes energéticos.
Por tanto, las hembras se beneficiarán más dispersándose y eligiendo entre estos machos territoriales.
Además, la hipótesis de Edipo también afirma que los padres en sistemas poligínicos desalojarán a los hijos con potencial para cornearlos.
[12] La supresión reproductiva ocurre cuando a los individuos sexualmente maduros de un grupo se les impide reproducirse debido a estímulos conductuales o químicos de otros miembros del grupo que suprimen el comportamiento reproductivo.
Si las condiciones actuales para la reproducción son desfavorables, como cuando sólo se presenta la endogamia como medio para reproducirse, los individuos pueden aumentar su éxito reproductivo a lo largo de su vida programando sus intentos reproductivos para que ocurran durante condiciones más favorables.
(2000) examinó los grupos sociales de suricatas y los factores que afectan a la supresión reproductiva en las hembras subordinadas.
La reproducción requería tanto la presencia de una pareja no emparentada del sexo opuesto, que actuaba como estímulo apropiado sobre los subordinados suprimidos reproductivamente que estaban quiescentes en presencia del individuo dominante original.
Para las hembras, las cópulas extra-pareja aseguran la fertilización de los huevos y proporcionan una mayor variedad genética con espermatozoides compatibles que evitan la expresión de genes recesivos dañinos que vienen con la endogamia.
En las cópulas extra-pareja, tanto los costes de la endogamia como la pérdida de la pareja masculina (que lleva a la pérdida del cuidado paterno) deben considerarse con los beneficios del éxito reproductivo que proporciona la cópula extra-pareja.
Cuando el cuidado paterno está ausente o tiene poca influencia en la supervivencia de la descendencia, suele ser favorable para las hembras realizar cópulas extrapareja para aumentar el éxito reproductivo y evitar la endogamia.
La mayor parte de la bibliografía sobre la evitación de la endogamia se publicó hace al menos 15 años, lo que ha permitido el crecimiento y desarrollo del estudio mediante métodos experimentales y tecnología contemporáneos.