Motilidad bacteriana
Del mismo modo, las bacterias magnetotácticas alinean su movimiento con el campo magnético de la Tierra.
[2] Einstein no sólo dedujo la difusión de partículas en suspensión en líquidos en reposo, sino que también sugirió que estos descubrimientos podían utilizarse para determinar el tamaño de las partículas; en cierto sentido, fue el primer microrreólogo del mundo.
[3] El número Reynolds, llamado así en su honor, cuantifica esta comparación como un cociente adimensional de las fuerzas inerciales y viscosas características:
[3][4] Como ilustración concreta, considere una vieira matemática que consta de dos piezas rígidas unidas por una bisagra.
Aquí se originó la sorprendente cita: «Rápida o lenta, recorre exactamente su trayectoria y vuelve al punto de partida".
[3] Este artículo sigue inspirando debates científicos en curso; por ejemplo, trabajos recientes del grupo Fischer, del Instituto Max Planck de Sistemas Inteligentes, confirmaron experimentalmente que el principio de la vieira sólo es válido para los fluidos newtonianos.
[3][6] Algunos de los sistemas móviles más pequeños que se conocen son las proteínas motoras, es decir, proteínas y complejos proteicos presentes en las células que llevan a cabo diversas funciones fisiológicas mediante la transducción de energía química en energía mecánica.
En las bacterias grampositivas, la membrana citoplasmática sólo está rodeada por una gruesa pared celular de peptidoglicano.
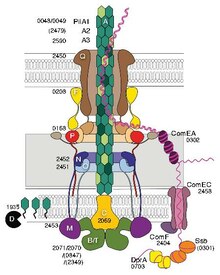
Los flagelos procariotas utilizan un motor rotatorio y los eucariotas un complejo sistema de filamentos deslizantes.
Son frágiles y se sustituyen constantemente, a veces con pili de diferente composición.
[3][53]El arquetipo de la natación bacteriana está representado por el organismo modelo Escherichia coli, muy estudiado.
La rotación en sentido horario desarma el haz y la célula gira al azar (tumbling).
[54][58] Rhodobacter sphaeroides, con su flagelación subpolar monótona, representa otra estrategia de motilidad:[24][55] el flagelo sólo gira en una dirección, y se detiene y se enrolla contra el cuerpo celular de vez en cuando, lo que provoca reorientaciones del cuerpo celular.
[56][59][60] En la bacteria del suelo Pseudomonas putida, un penacho de flagelos helicoidales está unido a su polo posterior.
En ese caso, el haz flagelar adopta la forma de una hélice zurda que gira en dirección CW, y las trayectorias son predominantemente rectas.
Sin cambiar su orientación, el cuerpo celular retrocede a través del haz aflojado.
Se pueden distinguir distintos tipos de taxia según la naturaleza del estímulo que controla el movimiento dirigido, como la quimiotaxis (gradientes químicos como la glucosa), la aerotaxis (oxígeno), la fototaxis (luz), la termotaxis (calor) y la magnetotaxis (campos magnéticos).
Evaluando repetidamente su rumbo y ajustándolo si se mueven en la dirección equivocada, las bacterias pueden dirigir su movimiento de paseo aleatorio hacia lugares favorables.
Si la bacteria detecta que se mueve en la dirección correcta (hacia el atrayente/lejos del repelente), seguirá nadando en línea recta durante más tiempo antes de dar tumbos; sin embargo, si se mueve en la dirección equivocada, dará tumbos antes.
[95] La naturaleza helicoidal del filamento flagelar individual es fundamental para que se produzca este movimiento.
Los vertebrados parecen haber aprovechado este hecho al poseer un receptor inmunitario (TLR5) diseñado para reconocer esta proteína conservada.
Muchas bacterias, como Vibrio, son monoflageladas y tienen un solo flagelo en un polo de la célula.
Aunque algunos trabajos han examinado perfiles químicos dependientes del tiempo,[103] las investigaciones anteriores sobre quimiotaxis con E. coli y otros organismos modelo han examinado habitualmente gradientes químicos estables lo bastante fuertes como para provocar una respuesta quimiotáctica perceptible.
Sin embargo, detectar tales gradientes a distancia tiene un enorme valor, porque apuntan hacia las fuentes de nutrientes.
Los gradientes poco profundos son importantes precisamente porque pueden usarse para navegar hasta regiones cercanas a las fuentes donde los gradientes se vuelven pronunciados, las concentraciones son altas y las bacterias pueden adquirir recursos a un ritmo elevado.
Es análoga a la quimiotaxis positiva, salvo que el atrayente es la luz y no una sustancia química.
Algunos procariotas filamentosos deslizantes pueden incluso percibir la dirección de la luz y realizar giros dirigidos, pero su movimiento fototáctico es muy lento.
[114][115] La señalización descendente en arquebacterias fototácticas implica CheA, una histidina quinasa, que fosforila el regulador de respuesta, CheY.
[128]Para realizar esta tarea, estas bacterias tienen orgánulos biomineralizados llamados magnetosomas que contienen cristales magnéticos.
[129] Las reacciones de escape no están estrictamente orientadas, sino que suelen implicar un movimiento hacia atrás, a veces con un componente geotáctico negativo.